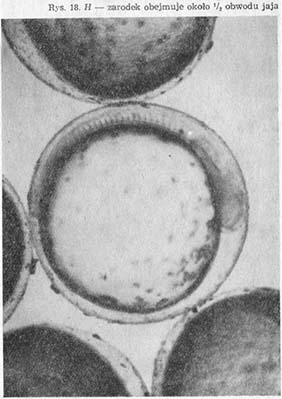
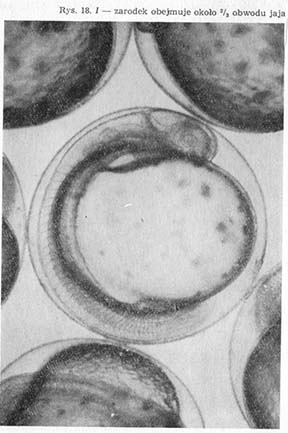
Procesowi temu towarzyszy zwiększanie się liczby miomerów oraz długości zarodka (rys. 18—H, I), który w stadium poprzedzającym wylęg nie mieści się już na obwodzie kuli żółtkowej.
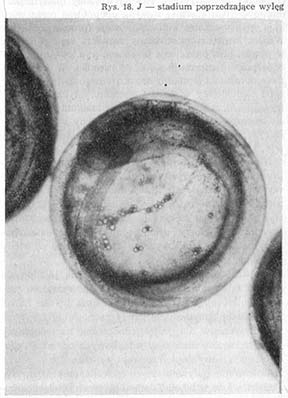
Przednia część głowy i ogon, sięgający końcem daleko poza tylną krawędź oka (rys. 18—J), oddzielone są wtedy od żółtka.
Szybkość rozwoju zarodkowego zależna jest przede wszystkim od temperatury. Przy jej podwyższeniu proces trwa krócej, przy obniżeniu przedłuża się. Ale nawet w tej samej temperaturze i przy zachowaniu identycznych warunków dla grupy jednocześnie zapłodnionych jaj poszczególne zarodki w jednakowych odcinkach czasu osiągają różny stopień rozwoju, co wskazuje na pewne zróżnicowanie indywidualnych cech komórek płciowych. Przedstawione w tabeli 7 dane o okresach dzielących kolejne stadia rozwoju dotyczą zarodków, które w temperaturze 10° rozwijały się najszybciej.
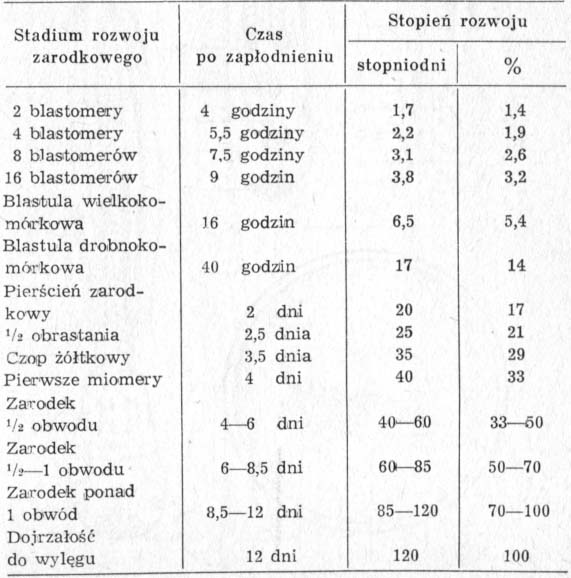 TABELA 7. Szybkość rozwoju zarodkowego szczupaka w temperaturze 10°C (według Lindrotha, 1946)
TABELA 7. Szybkość rozwoju zarodkowego szczupaka w temperaturze 10°C (według Lindrotha, 1946)
Zdaniem Lindrotha (1946), za ostatnie stadium rozwoju ikry należy uznać taki stan uformowania zarodka, w którym staje się on zdolny do wylęgu. Stadium to nie jest jednak równoznaczne z samym momentem wylęgania. Oprócz określonego progu dojrzałości muszą być spełnione dwa dalsze warunki: aktywność zarodka i dostateczne osłabienie wytrzymałości błony jajowej. Na głowie, a w mniejszej liczbie także na woreczku żółtkowym i tułowiu, są rozmieszczone tak zwane gruczoły wykluwania wydzielające substancję, która działając na błonę powoduje znaczne zmniejszenie jej grubości. Wzrastająca aktywność ruchowa przyczynia się do bardziej równomiernego rozprowadzenia wydzielonej substancji w całej wolnej przestrzeni.
Pierwsze ruchy w zapłodnionym jaju można obserwować już po dwóch, trzech godzinach. Polegają one na zmianie kształtu ii położenia kuli żółtkowej. Samodzielne ruchy zarodka pojawiają się z chwilą, gdy jego długość przekroczy połowę obwodu jaja. W pierwszej fazie są to słabe i sporadyczne wygięcia tułowia, później silniejsze ruchy ogona, zmieniające położenie całego ciała wraz z woreczkiem żółtkowym. W okresie poprzedzającym wylęg częstotliwość skurczów mięśniowych zwiększa się. Występują one seriami i wprawiają ciało zarodka w charakterystyczne drżenie.
Znaczenie ruchów zarodka podkreśla Kotlarewiskaja (1969), która obserwowała wylęg przy zróżnicowanym stanie wytrzymałości błon jajowych. Najaktywniejsze zarodki potrafiły wydostać się z jaj, gdy grubość błony przekraczała 8 μ, najmniej aktywne — dopiero przy grubości 1 μ. Czynnikiem wzmagającym ruchliwość jest temperatura. Rezniczenko (1958) podaje, że w ciągu 10 minut zarodki wykonywały 1—2 ruchy przy 5°, 5—6 przy 15° i 14—15 ruchów przy 25°. Aktywizujący wpływ oświetlenia przedstawiono na rysunku 20.
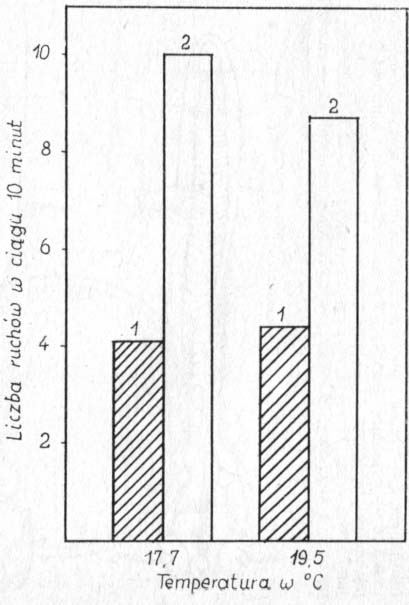 Rys. 20. Aktywność ruchowa zarodków szczupaka w zależności od oświetlenia: 1 — przy zaciemnieniu (16 luksów), 2 — w pełnym świetle słonecznym (1000—1200 luksów) (wg Kotlarewskiej).
Rys. 20. Aktywność ruchowa zarodków szczupaka w zależności od oświetlenia: 1 — przy zaciemnieniu (16 luksów), 2 — w pełnym świetle słonecznym (1000—1200 luksów) (wg Kotlarewskiej).








