Skład pokarmu szczupaka jest więc wypadkową wzajemnego układu elementów środowiskowych i populacyjnych, a najważniejszą rolę w jego kształtowaniu odgrywa aktualna liczebność najbardziej dostępnych ofiar. Sformułowanie to uwzględnia różnice spowodowane odmiennym składem ichtiofauny w odległych od siebie regionach geograficznych, w poszczególnych zbiornikach wodnych jednego rejonu, a także na odrębnych stanowiskach tego samego zbiornika; uwzględnia ono również zmiany sezonowe, które następują w cyklu rocznym oraz zmiany zachodzące z wiekiem drapieżników. Dokładną charakterystykę pokarmu należałoby oprzeć na szczegółowych analizach w obrębie wąskich nisz ekologicznych. Brak miejsca nie pozwala na wyczerpujące przedstawienie bogato prezentowanych w literaturze ichtiologicznej materiałów tego typu. Zaczerpnięte z badań Antosiak (1963) zbiorcze dane dla dwudziestu zróżnicowanych limnologicznie jezior węgorzewskich obrazują stan najczęściej spotykany w wodach stojących północnej Europy.
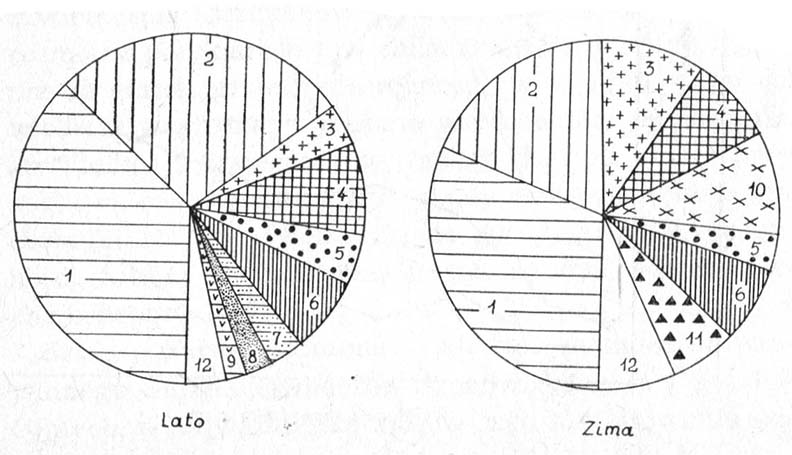 Rysunek. Skład pokarmu szczupaka z jezior węgorzewskich według częstości występowania: 1 — płoć, 2 — okoń, 3 — ukleja, 4 — krąp, 5 — ciernik, 6 — jazgarz, 7 — karaś, 8 — wzdręga, 9 — kiełb, 10 — leszcz, 11 — stynka, 12 — inne gatunki (wg Antosiak).
Rysunek. Skład pokarmu szczupaka z jezior węgorzewskich według częstości występowania: 1 — płoć, 2 — okoń, 3 — ukleja, 4 — krąp, 5 — ciernik, 6 — jazgarz, 7 — karaś, 8 — wzdręga, 9 — kiełb, 10 — leszcz, 11 — stynka, 12 — inne gatunki (wg Antosiak).
Wyraźna przewaga częstości występowania płoci i okonia świadczy o szczególnie wysokiej dostępności obu gatunków. Dominują one także w pokarmie szczupaka z tych jezior, w których inne ryby litoralowe nie ustępują im liczebnością. Listę pospolicie notowanych składników pokarmu uzupełniają: krąp, jazgarz, ukleja. Lokalne znaczenie, istotne w pewnych tylko niszach środowiskowych, mogą mieć: wzdręga, karaś, karp, kiełb, a nawet ciernik, dla którego ostre kolce płetw grzbietowej i brzusznych nie stanowią dostatecznej ochrony. Natomiast sielawa, stynka i leszcz należą do grupy ofiar podlegających presji okresowej — zwykle w czasie tarła lub zimowania. Sporadycznie w przewodach pokarmowych drapieżników spotyka się wszystkie pozostałe gatunki ryb zasiedlających dany zbiornik.
Kanibalizm wśród dorosłych szczupaków notowany jest częściej niż w okresie larwalnym ze względu na znaczniejsze zróżnicowanie wielkości osobników. Mimo to na ogół nasilenie jego nie wykracza poza przypadki sporadyczne, gdyż dostępność szczupaka jako ofiary ograniczona jest z jednej strony małą jego ruchliwością, z drugiej zaś szybkim wzrostem, który pozwala w krótkim czasie osiągnąć rozmiary chroniące przed presją własnego gatunku, skierowaną przede wszystkim na pogłowie drobnych ryb. Przy dużym zagęszczeniu populacji może jednak zjawisko kanibalizmu przybrać ostrzejsze formy i dlatego uważane jest za jeden z przejawów biologicznej samoregulacji liczebności drapieżnika. Stwarza ono również szansę utrzymania populacji przy zupełnym braku innych ryb w zbiorniku. Starsze osobniki, żywiąc się własnym potomstwem, mogą wtedy pośrednio wykorzystywać niedostępne dla nich zasoby pokarmu złożonego z fauny bezkręgowej (Kluczariewa, 1956).
Przysłowiowa żarłoczność szczupaków znajdowała wyraz w mocno przesadzonych liczbach, jakimi dawniej określano ilość zużywanego przez nie pokarmu. Według powszechnie panujących wówczas poglądów, przyrost drapieżnika o 1 kg miał kosztować kilkadziesiąt kilogramów zjedzonych ryb. Dopiero eksperymenty Scholza (1932), potwierdzone później przez innych autorów (Karzinkin, 1955; Johnson, 1966), dowiodły, że współczynnik pokarmowy — wskazujący ile jednostek ciężaru pokarmu potrzeba na wytworzenie jednostki ciężaru ciała — jest dla szczupaka karmionego rybami niższy niż dla innych ryb drapieżnych i wynosi 3—4 w okresie poprzedzającym dojrzałość płciową. Ekonomia wykorzystania pokarmu zależy w znacznej mierze od tego, jak wielka jego część zostaje zużyta dla samego tylko utrzymania organizmu przy życiu — to znaczy na pokrycie strat energii wydatkowanej na pracę mięśni w czasie oddychania d ruchu oraz na przetrwanie już wytworzonych tkanek. Długie okresy osłabionej aktywności w czasie wyczekiwania na zdobycz stawiają szczupaka w korzystnej sytuacji. Według badań Johnsona (1966), tygodniowa dawka pokarmu potrzebnego na pokrycie strat energetycznych wynosi zaledwie 20—50 mg na 1 g ciężaru ciała i wszystko, oo zdoła drapieżnik zjeść ponad tę ilość, zamienia się w nowe tkanki, czyli zostaje zużyte na wzrost. Osobniki niedojrzałe płciowo mogą na ten cel zużywać 70% całej przyjmowanej porcji pokarmowej, później jednak zapotrzebowanie na składniki odżywcze wzrasta w związku z produkcją corocznie wydalanych z organizmu komórek rozrodczych. Zostaje ono pokryte kosztem składników zużywanych dotychczas na przyrastanie ciała, których ilość, w populacjach złożonych z osobników o (zróżnicowanym wieku, zmniejsza się — według danych Johnsona — do 20 % w stosunku do całego zjadanego pokarmu. Efektem jest wyraźne pogorszenie się współczynnika pokarmowego. Dalsze obliczenia cytowanego autora (Johnson, 1966) wykazały, że w badanym przez niego jeziorze Windermere spożycie potrzebne na przyrost 1 kg szczupaka wynosi około 10 kg pokarmu. Ponieważ średni roczny przyrost ciężaru jednego osobnika tej populacji jest mniejszy, ilość ryb zjadanych w ciągu roku wynosi przeciętnie 3—4 kg na 1 kg żywej masy drapieżników zasiedlających zbiornik.








