Presja pokarmowa młodych szczupaków na pokolenie płoci z tego samego rocznika trwa nadal po zakończeniu rozwoju larwalnego obydwu gatunków i po ich rozproszeniu się wzdłuż strefy litoralowej. Obok płoci pojawia się wtedy w pokarmie również najmłodszy rocznik okonia, a później, w miarę szybkiego wzrostu drapieżników, starsze roczniki drobnych ryb (ukleja, jazgarz, krąp). Bezkręgowce, w tym również plankton, stają się pokarmem okolicznościowym, zjadanym przypadkowo lub — przy ubogim rybostanie — przymusowym. Konieczność żywienia się fauną bezkręgową, nawet w następnych latach życia, jest częściej udziałem szczupaków z wód płynących niż z jezior. Na przykład w duńskich rzekach pstrągowych Larsen (1951) stwierdził znaczne ilości skorupiaków, robaków i larw owadów w żołądkach szczupaków osiągających długość do 40 cm, a mniej licznie występowały nawet u większych.
W celu zilustrowania siły „instynktu” drapieżnictwa warto raz jeszcze powołać się na wspomniany wcześniej eksperyment Gimmy’ego (1953). Młody szczupak (około 5 cm długości), oddzielony szklaną szybą od ofiary, rzucał się na nią w pierwszej fazie doświadczenia 260 razy w ciągu godziny, mimo że każdy skok kończył się silnym zderzeniem z przegrodą. W miarę upływu czasu częstotliwość ataków malała, lecz zamierzony cel eksperymentu — zupełny brak zainteresowania obiektem — osiągnięto dopiero po 140 dniach od rozpoczęcia tresury. Ćwiczone w ten sam sposób kilkunastocentymetrowe okonie rezygnowały z podejmowania ataków już po 22 dniach nauki. Nic więc dziwnego, że i w warunkach naturalnych dorosłe szczupaki rzucają się bez wyboru na wszystko, co wokół nich się rusza lub jest poruszane. Aby je do tego sprowokować, nie zawsze konieczne jest imitowanie kształtu ryby za pomocą blaszanych błystek spinningowych. Autorzy popularnych opracowań ichtiologicznych i wędkarskich prześcigają się w wyliczaniu najdziwniejszych przypadków. Obserwowano np. szczupaka atakującego unoszone falą pudełko po papierosach (Aldinger, 1965), a w przewodach pokarmowych znajdowano takie przedmioty, jak łyżka, nóż, widelec, zegarek kieszonkowy z łańcuszkiem czy nawet ostrą amunicję karabinową (Hegemann, 1964). Przykłady te dowodzą, że u dorosłych szczupaków trudno byłoby doszukiwać się oznak jakiejkolwiek wybiórczości pokarmowej. Jedynym kryterium wyboru jest ruch, za pomocą którego przedmiot sygnalizuje swoją obecność, a dodatkowym warunkiem jego wielkość, nadająca sygnałom odpowiednie natężenie. Doświadczenia wędkarzy wykazują, że w okresach mniej intensywnego żeru drapieżniki zwracają uwagę także na kształt, barwę, sposób i szybkość poruszania się obserwowanego obiektu.
Chwytając wszystko, co się porusza, szczupak nie omija żab, myszy oraz innych drobnych ssaków przypadkowo trafiających do wody, a także — o czym wspomniał wcześniej cytowany Krzysztof Kluk — młodych ptaków pływających. Znakomitą większość obiektów poruszających się w wodzie i mających odpowiednie rozmiary stanowią jednak ryby.
Jeśliby wszystkie gatunki ryb żyjące w zbiorniku wodnym mogły być w równym stopniu potencjalnymi ofiarami szczupaków, wówczas należałoby oczekiwać, że skład pokarmu tych ostatnich będzie wiernym odzwierciedleniem liczebności składników rybostanu. Jednakże sytuacje takie zdarzają się tylko wyjątkowo. Istnieje szereg czynników, które różnicując stopień dostępności poszczególnych ofiar, stwarzają odmienne typy biologicznych powiązań między nimi a drapieżnikiem. Jednym z najważniejszych jest zbieżność stref środowiskowych. Ryby pelagiczne, przydenne i prądolubne mniej są narażone na presję ze strony szczupaka niż gatunki stale współbytujące z nim w obrębie litoralu. Nie wszystkie jednak są całkowicie eliminowane z jego pokarmu, ponieważ cechy biologiczne obu stron mogą doprowadzać do okresowych zbliżeń w zasięgu występowania. Szczególnie krytycznym dla wielu gatunków staje się okres ich rozrodu, związany mniej lub bardziej ze strefą przybrzeżną i z tworzeniem znacznych skupień w zasięgu penetracji drapieżników. Toteż przede wszystkim z rozbieżności terminów tarła ryb słodkowodnych wynika często obserwowana zmienność składu pokarmu szczupaka w okresie od wiosny do jesieni. Inne warunki żerowiskowe mogą również wytworzyć się po ochłodzeniu wody, kiedy populacje niektórych gatunków, wykazując zmniejszoną ruchliwość, tworzą liczebne skupienia na ograniczonej przestrzeni zimowisk.
Dostępność gatunków wchodzących w skład zespołu współbytującego ze szczupakiem w strefie przybrzeżnej jest także zróżnicowana ich właściwościami biologicznymi. Ważną rolę odgrywa tu wężej pojmowane siedlisko. Na przykład ryby związane z rejonami gęsto porośniętymi znajdują kryjówki chroniące przed atakiem. Drugim czynnikiem jest rozkład aktywności ofiar w cyklu dobowym. Gatunki żerujące nocą, a w ciągu dnia przejawiające niewielką aktywność, rzadziej zagrożone są spotkaniem z nieruchomo czatującymi drapieżnikami. Wreszcie zdolność szybkiego pływania, w połączeniu ze zwrotnością, stwarza niektórym ofiarom większe szanse ucieczki.
Obok właściwości biologicznych na dostępność ofiar wpływają także ich cechy morfologiczne — mianowicie rozmiary i kształt ciała — oraz wielkość atakujących je szczupaków. Choć z wiekiem, w miarę zwiększania się pyska i gardzieli, drapieżnik zdolny jest połykać coraz to większe ryby, sposobność po temu nie zdarza mu się zbyt często, gdyż występują one na ogół w rozproszeniu, a przy tym sprawnością pływania przewyższają drobnicę. Natomiast dostępność drobnych ryb praktycznie nie ulega zmianom, ponieważ główny czynnik ograniczający, jakim w tym przypadku może być mniejszy zasięg postrzegania jednej ofiary, jest neutralizowany właściwą drobnicy dążnością do tworzenia stad, których przemieszczenia wywołują zwielokrotnione falowanie wody (Domaniewskij, 1961). Toteż zestawienie wielkości ofiar z wielkością szczupaków wskazuje, że średnie rozmiary połykanych ryb zbliżone są bardziej do dolnej niż do górnej granicy dostępności, w wyniku czego ich wielkość względna, wyrażona w procentach w stosunku do wielkości drapieżników, maleje.
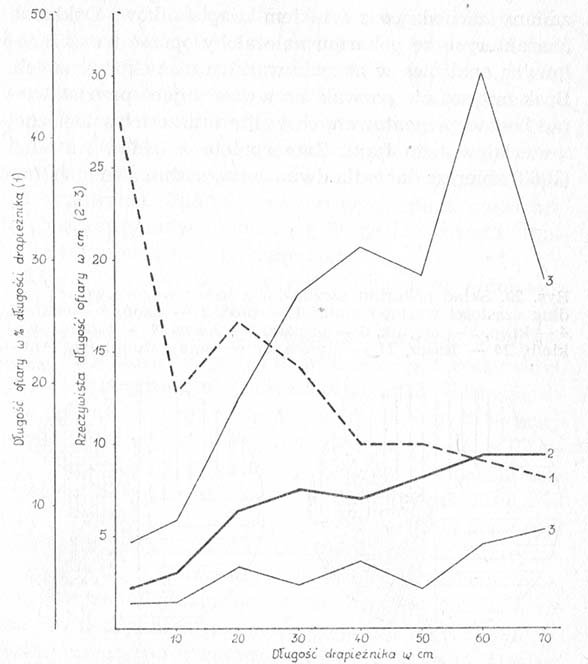 Rysunek. Zależność między długością ofiar a długością szczupaków z delty Wołgi: 1 — długość względna ofiar, 2 — średnia długość rzeczywista, 3 — wartości skrajne (wg Popowej).
Rysunek. Zależność między długością ofiar a długością szczupaków z delty Wołgi: 1 — długość względna ofiar, 2 — średnia długość rzeczywista, 3 — wartości skrajne (wg Popowej).
Kształt ciała ofiary odgrywa rolę zwłaszcza wtedy, gdy atakowane mogą być osobniki zbliżone wielkością do górnej granicy dostępności. Silniej wygrzbiecona forma stanowi wówczas cechę ochronną.








