Pokarm i odżywianie się szczupaków
Rodzaj pobieranego pokarmu stanowi podstawę podziału ryb na gatunki „spokojnego żeru” i drapieżne. Szczupak zaliczany jest do najbardziej typowych drapieżników. Oznacza to, że w ciągu prawie całego życia odżywia się niemal wyłącznie rybami i innymi kręgowcami oraz że tylko ten rodzaj pokarmu może spełniać warunek pełnego zaspokojenia fizjologicznych i biologicznych potrzeb organizmu.
W pierwszej fazie rozwoju jedynym źródłem pokarmu jest zawartość woreczka żółtkowego. Ale jeszcze przed całkowitym jej zresorbowaniem, po upływie 6—8 dni od wyklucia się, osiągając długość 10—12 mm, larwy zaczynają chwytać najmniejsze formy zooplanktonu. Okres, W którym drobny plankton jest głównym składnikiem pokarmu, nie przekracza jednego tygodnia. W tym czasie przewód pokarmowy — pierwotnie ukształtowany w formę prostej rurki — różnicuje się wytwarzając pętlę jelita i żołądek.
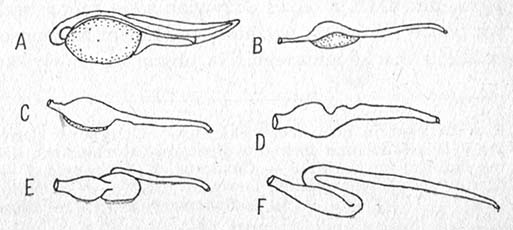 Rysunek. Budowa przewodu pokarmowego larw szczupaka o długości: A — 8,0 mm, B — 10,2 mm, C — 12,2 mm, D — 13 mm, E — 14,7 mm, F — 17,5 mm (wg Frost).
Rysunek. Budowa przewodu pokarmowego larw szczupaka o długości: A — 8,0 mm, B — 10,2 mm, C — 12,2 mm, D — 13 mm, E — 14,7 mm, F — 17,5 mm (wg Frost).
Z chwilą wyodrębnienia się żołądka — przy długości 15—17 mm — następuje istotna zmiana sposobu odżywiania się. Żerujące larwy, w miarę dalszego wzrostu, dążą do wybierania coraz większych organizmów, co jest uzasadnione koniecznością ekonomicznego wydatkowania energii (Karzinkin, 1955). Technika żerowania bowiem, od samego początku — podobnie jak u osobników dorosłych — polega na oddzielnym atakowaniu każdego obiektu. Przy niezmienionych rozmiarach ofiar, a stale wzrastającej masie zużywanego pokarmu, liczba podejmowanych ataków rychło musiałaby przekroczyć możliwości organizmu. Toteż drobne wioślarki i widłonogi stopniowo zastępowane są największymi formami planktonu, larwami owadów i ryb.
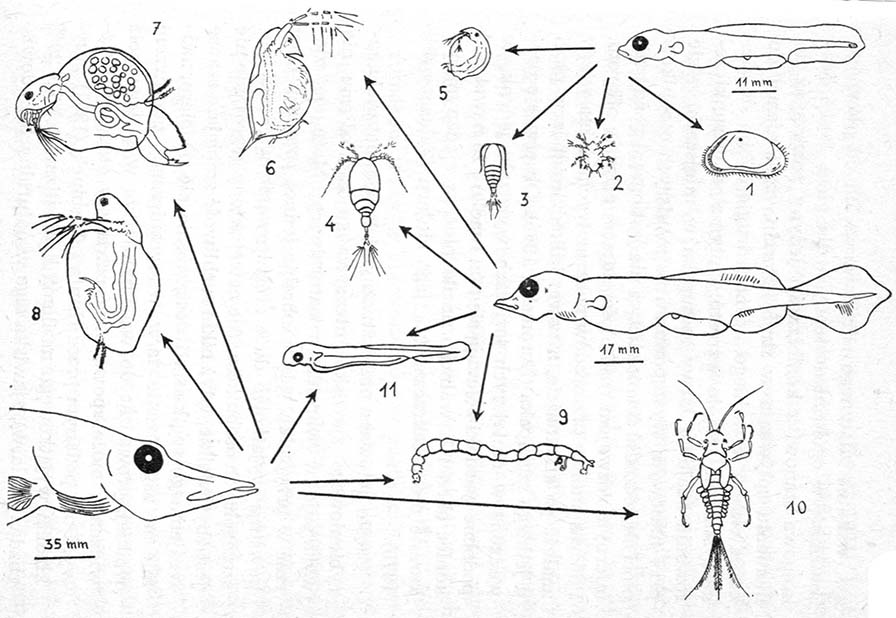 Rysunek. Pokarm larw szczupaka: 1 — małżoraczek (Ostracoda), 2 i 3 — młodociane formy widłonogów (Cyclopidae), 4 — forma dorosła; wioślarki: 5 — Chydorus, 6 — Daphnia, 7 — Eurycercus, 8 — Sirnocephalus; larwy owadów: 9 — Tendipedidae, 10 — Ephemeroptera; 11 — larwa płoci.
Rysunek. Pokarm larw szczupaka: 1 — małżoraczek (Ostracoda), 2 i 3 — młodociane formy widłonogów (Cyclopidae), 4 — forma dorosła; wioślarki: 5 — Chydorus, 6 — Daphnia, 7 — Eurycercus, 8 — Sirnocephalus; larwy owadów: 9 — Tendipedidae, 10 — Ephemeroptera; 11 — larwa płoci.
Procesowi temu sprzyjają przeobrażenia budowy ciała, zwłaszcza zwiększanie się pyska i rozwój zębów gardzielowych, służących do przepychania pokarmu ku przełykowi. Natomiast mechanizmem wyzwalającym nowy typ reakcji na poruszenia dużych obiektów w sąsiedztwie stanowiska zajmowanego przez larwę staje się narząd linii bocznej, który w pierwszej fazie jej żerowania — przy minimalnym zasięgu wzroku — pełnił funkcję ostrzegawczą, związaną z odruchem ucieczki.
Moment uformowania się żołądka oznacza gotowość podjęcia drapieżnictwa. W sprzyjających warunkach może to nastąpić w niecałe dwa tygodnie po wylęgu i odtąd larwy ryb stają się dla młodych szczupaków pokarmem najkorzystniejszym. Nie tylko duża koncentracja masy pokarmowej w jednym obiekcie, ale również większa ilość substancji dającej się strawić i przyswoić stanowi o ich wysokiej — w porównaniu z fauną bezkręgową — wartości odżywczej. W większości naszych jezior, zwłaszcza wielkich, rejon przybrzeżnych tarlisk szczupaka jest wykorzystywany przez później od niego trącą się płoć. Różnica w terminie tarła obu gatunków doprowadza zazwyczaj do sytuacji, w której masowo wylęgające się larwy płoci wchodzą od razu w spektr pokarmowy czekających na nie młodych drapieżników. Często jednak warunki termiczne, opóźniające tarło i wylęg płoci, przedłużają okres odżywiania się fauną bezkręgową ponad minimum wyznaczone stanem zaawansowania rozwoju morfologicznego. Na obszarach zalewiskowych zaś, wskutek znacznego odizolowania ich od głównego zbiornika, w ogóle niedostępnych dla płoci, fauna bezkręgowa stanowi z konieczności jedyny pokarm aż do momentu spływania w strefę litoralu, czyli niemal do końca okresu larwalnego. Jedno i drugie odbija się niekorzystnie na tempie wzrostu. Optymalne warunki żerowania znajdują larwy szczupaka w miejscach, gdzie przy masowym występowaniu najmłodszych osobników płoci skuteczność drapieżnictwa nie jest ograniczona nadmiernym rozwojem roślinności dającej ochronę ofiarom. Podczas analizy przewodów pokarmowych można wtedy znaleźć po kilkanaście, a nawet kilkadziesiąt larw płoci u jednego szczupaka (Żuromska, 1966). Obserwacje prowadzone w mniejszych jeziorach wykazały jednak, że podobne warunki zdarzają się tam rzadko i że nawet pod koniec okresu larwalnego udział płoci w zjadanym pokarmie nie przekracza 40% jego ciężaru (Załachowski, 1970).
Często notowane przy sztucznym chowie materiału zarybieniowego zjawisko intensywnego kanibalizmu nie występuje w warunkach naturalnych. Może się zdarzać w przypadku, gdy w obrębie jednej populacji larw powstaje wyjątkowo duże zróżnicowanie wielkości poszczególnych osobników. Ale nawet wtedy zagrożenie nie jest zbyt wielkie, gdyż nikła ruchliwość zmniejsza szanse wzajemnego spotkania i dostrzeżenia się. Dopiero nadmierne zagęszczenie larw w ograniczonej przestrzeni wodnej, prowadząc do współbytowania w zasięgu pola widzenia, może spowodować ostrzejszy stan konfliktowy. Poza podanym przez Hunta i Carbine’a (1951) przykładem wyjątkowego nasilenia kanibalizmu (około 20% zbadanych larw), zjawiska tego w naturalnych środowiskach bądź w ogóle nie stwierdzono (Frost, 1954), bądź notowano sporadycznie (Makkowiejewa, 1956; Franklin, Smith, 1963; Załachowski, 1970 i inni).








