Rozmyślnie użyto tutaj określenia „połykana ryba”, ponieważ szczupak nie gryzie ani nie rozdrabnia pokarmu znajdującego się w pysku. Dlatego niepotrzebne mu są duże zęby na kościach szczęki górnej i tylko przednia jej część (kość przedszczękowa) zaopatrzona jest w szereg niewielkich zębów chwytnych.
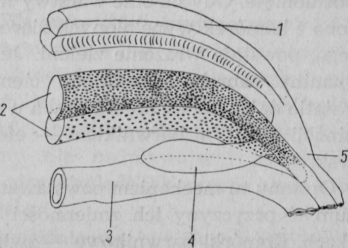
Pochwycona ofiara może być przełknięta pod warunkiem, że zostanie skierowana do gardzieli głową. Toteż szczupak chwyciwszy w poprzek — co zdarza się często — lub z tyłu, zmuszony jest, manewrując szczękami i językiem, odpowiednio ją ustawić (rys. 7).
 Rys. 7. Szczupak obracający pochwyconą rybę (reprodukcję z książki „The Book of Fishes“, Washington 1958, wykonał C. Nagięć)
Rys. 7. Szczupak obracający pochwyconą rybę (reprodukcję z książki „The Book of Fishes“, Washington 1958, wykonał C. Nagięć)
Czyni to na ogół zadziwiająco sprawnie, choć walka z dużą rybą może trwać nawet kilkanaście minut. Obserwowano przy tym krótkie, urywane wyrzuty głowy w bok, skąd wracała do pozycji wyjściowej nie odchylając się w przeciwną stronę (Hoogland i inni, 1957). Wykonywane jednocześnie ruchy szczękami, podobne do gryzienia lub żucia, mają na celu przesunięcie ofiary ku przełykowi.
Z reguły nie spotyka się szczupaków, które by miały pełny komplet zębów chwytnych na dolnej szczęce. Wiąże się to z mechanizmem ich wymiany, trwającej nieprzerwanie w ciągu całego życia. W sąsiedztwie każdego zęba roboczego, pod poduszeczką z miękkich tkanek, tkwi zawsze kilka zawiązków zębowych gotowych go zastąpić. Miejsce ich osadzenia znaczą brązowawe plamy lub zaczerwienione wzgórki. Z chwilą, gdy „stary” ząb wychodzi z użycia i zostaje zresorbowany przez narastającą tkankę, na jego miejsce sadowi się najbliżej leżący zawiązek. W okresie wzrostu trzon jego jest miękki, a umocowanie jeszcze niepewne, tak że łatwo daje się odchylić we wszystkie strony. Odróżnić młody ząb można także po tym, że jest skierowany ostrzem do wnętrza jamy gębowej. Proces wymiany trwa nieprzerwanie, ponieważ robocze zęby nigdy nie ustępują jednocześnie i obok nich widnieją na szczęce dolnej nowe w różnych fazach rozwoju. Tłumaczenie sezonowych zmian intensywności żerowania szczupaków wymianą zębów jest wobec tego nie uzasadnione, chyba że udowodniono by zależne od pory roku różnice w nasileniu procesu wymiany (Barsukow, 1962). Zupełny lub prawie zupełny brak zębów chwytnych przypisać należy zmianom patologicznym. W takich przypadkach nie ma również zawiązków zębów do wymiany.
Ponieważ połykana ryba przedostaje się do żołądka w całości, narząd ten, podobnie jak przełyk, musi mieć odpowiednie rozmiary. Tymczasem żołądek szczupaka, kształtem podobny do wrzeciona, jest stosunkowo wąski, przełyk zaś ma w przekroju średnicę nie większą niż 2 cm. Jednakże grube ściany, od wewnątrz ścieśnione w kilkanaście podłużnych fałd, zanikających przy zetknięciu się z pokarmem, nadają obydwu tym częściom przewodu pokarmowego niezwykłą rozciągliwość. Z wąskiego, wrzecionowatego żołądka powstaje wtedy pojemny wiórek o cienkich, półprzezroczystych ściankach. Gdy zdobycz mimo to nie mieści się, trawienie może przebiegać stopniowo, przy czym często ogon połkniętej ryby wystaje wtedy z gardzieli czekając na swoją kolej.
Przełyk przechodzi w żołądek bez wyraźnie widocznego zróżnicowania struktury. Granicę obu części stanowi początek strefy gruczołów wydzielających do żołądka enzymy trawienne. Jednym z mich jest pepsyna — enzym uczestniczący w rozkładzie substancji białkowych. Działa on tylko w środowisku kwaśnym, toteż odczyn we wnętrzu żołądka wynosi podczas trawienia 4,5—4,7 pH. Badania, które przeprowadzili Vonk i Meninega (1938), wykazały, że tylko na samej powierzchni rozkładanego pokarmu pH obniża się do 2,4—3,6, co odpowiada w przybliżeniu wartościom notowanym u wyższych kręgowców i uważanym za odczyn aktywizujący pepsynę. Proces trawienia nie obejmuje więc jednocześnie całej masy pokarmowej, lecz stopniowo postępuje w głąb ku kolejno odkrywanym warstwom. W efekcie trawienie jednej porcji ciągnie się dość długo i trwa zazwyczaj 3—5 dni.
Połączenie żołądka z jelitem ścieśnione jest przez silnie umięśniony zwieracz odźwiernika, który ogranicza kontakt między środowiskami różniącymi się odczynem. Enzymy jelitowe, uczestniczące w dalszym rozkładzie rozdrobnionej „papki” pokarmowej, działają przy pH powyżej 7. Jelito, tworzące u szczupaka pojedynczą pętlę, dzieli się na część środkową i końcową, którą łatwo można odróżnić po ciemniejszym zabarwieniu. Jest ono, podobnie jak u innych ryb drapieżnych, stosunkowo krótkie i na ogół nie przewyższa 1,1—1,25 długości ciała (Hegemann, 1964). Część środkowa jest najdłuższym odcinkiem przewodu pokarmowego i stanowi ponad połowę całej jego długości.
Jelito kończy się odbytem umiejscowionym przed nasadą płetwy odbytowej. Tuż za nim (rys. 8) znajdują się dwa dalsze odrębne otwory: przez pierwszy wydalane są w czasie tarła produkty gruczołów płciowych, drugi odprowadza mocz z nerek.
 Rys. 8. Schemat zakończenia układu moczopłciowego i jelita szczupaka: 1 — nerka, 2 — jajniki, 3 — jelito końcowe, 4 — pęcherz moczowy, 5 — moczowód (wg Schillinga za Hegeimannem).
Rys. 8. Schemat zakończenia układu moczopłciowego i jelita szczupaka: 1 — nerka, 2 — jajniki, 3 — jelito końcowe, 4 — pęcherz moczowy, 5 — moczowód (wg Schillinga za Hegeimannem).
Niewielka, lecz dostrzegalna, różnica w kształcie otworu odbytowego jest u szczupaków jedyną cechą zewnętrzną, po której można odróżnić płeć już w pierwszym roku życia (Demczenko, 1963).
Budowa i działanie aparatu skrzelowego nie odbiega w zasadniczych punktach od wzoru spotykanego u większości ryb kostnoszkieletowych. Warto jednak zwrócić uwagę, że wielki pysk i szerokie szczeliny skrzelowe — zdolne przepuszczać w krótkim czasie pokaźny strumień wody — stwarzają szczupakowi dobre warunki oddechowe. Ponadto stosunkowo duża powierzchnia pokrytych drobnymi naczyniami krwionośnymi płatków skrzelowych (1,5 cm2 na 1 gram ciężaru ryby — według Hegemanna, 1964), a także łatwość wiązania tlenu przez hemoglobinę, która wysyca się przy stosunkowo niskich ciśnieniach parcjalnych, sprawia, że dorosłe szczupaki nieźle znoszą pobyt w wodach niezbyt bogatych w tlen i „duszności” zaczynają odczuwać dopiero wtedy gdy jego zawartość spadnie do 2—3 mg/l (Priwolniew, 1954.)








